DNA-dependent protein kinase (DNA-PK) is a nuclear protein Serine (Ser)/Threonine (Thr) kinase that acts as both a molecular sensor and transmitter of DNA damage, and plays important roles in the DNA repair of double stranded breaks (DSBs), mediating immunoglobulin V(D)J gene recombination events, as well as telomere stabilization.
-
 Australia
Australia
-
 Austria
Austria
-
 Belgium
Belgium
-
 Brazil
Brazil
-
 Canada
Canada
-
 China
China
-
 Czech Republic
Czech Republic
-
 Denmark
Denmark
-
 Finland
Finland
-
 France
France
-
 Germany
Germany
-
 Greece
Greece
-
 Hong Kong
Hong Kong
-
 Hungary
Hungary
-
 Iceland
Iceland
-
 India
India
-
 Ireland
Ireland
-
 Israel
Israel
-
 Italy
Italy
-
 Japan
Japan
-
 Korea
Korea
-
 Luxembourg
Luxembourg
-
 Malaysia
Malaysia
-
 Netherlands
Netherlands
-
 New Zealand
New Zealand
-
 Norway
Norway
-
 Poland
Poland
-
 Qatar
Qatar
-
 Romania
Romania
-
 Saudi Arabia
Saudi Arabia
-
 Singapore
Singapore
-
 Spain
Spain
-
 Sweden
Sweden
-
 Switzerland
Switzerland
-
 Taiwan
Taiwan
-
 Turkey
Turkey
-
 United Kingdom
United Kingdom
-
 United States
United States
DNA-PK Inhibitors
| Cat.No. | Product Name | Information | Product Use Citations | Product Validations |
|---|---|---|---|---|
| S8586 | Nedisertib (M3814) | Nedisertib (M3814, Peposertib, MSC2490484A) is an orally bioavailable, highly potent and selective inhibitor of DNA activated protein kinase (DNA-PK) with an IC50 of < 3 nM. |
|
|
| S8843 | AZD7648 | AZD7648 is a potent inhibitor of DNA-PK with an IC50 of 0.6 nM in biochemical assay and more than 100-fold selective against 396 other kinases. |
|
|
| S2638 | NU7441 (KU-57788) | NU7441 (KU-57788) is a highly potent and selective DNA-PK inhibitor with IC50 of 14 nM. It also inhibits mTOR and PI3K with IC50 of 1.7 μM and 5 μM in cell-free assays, respectively, and reduces the frequency of NHEJ while increasing the rate of HDR following Cas9-mediated DNA cleavage. |
|

|
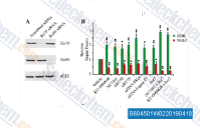
| S8045 | KU-0060648 | KU-0060648 is a dual inhibitor of DNA-PK and PI3Kα, PI3Kβ, PI3Kδ with IC50 of 8.6 nM and 4 nM, 0.5 nM, 0.1 nM respectively, less inhibition of PI3Kγ with IC50 of 0.59 μM. |
|
|
| S8593 | VX-984 | VX-984 (M9831) is an orally active, potent, selective, and ATP-competitive inhibitor of DNA-PK. This compound effectively suppresses non-homologous end joining (NHEJ) and increases DNA double-strand breaks (DSBs). It enhances the cytotoxic effects of ionizing radiation (IR) in various cancer cell lines, including non-small cell lung cancer (NSCLC) cell lines, in vitro. Additionally, this inhibitor reduces DNA-PKcs autophosphorylation. | ||
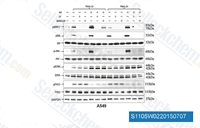
| S1105 | LY294002 | LY294002 (SF 1101, NSC 697286) is the first synthetic molecule known to inhibit PI3Kα/δ/β with IC50 of 0.5 μM/0.57 μM/0.97 μM, respectively; more stable in solution than Wortmannin, and also blocks autophagosome formation. It not only binds to class I PI3Ks and other PI3K-related kinases, but also to novel targets seemingly unrelated to the PI3K family. This compound also inhibits CK2 with IC50 of 98 nM. It is a non-specific DNA-PKcs inhibitor and activates autophagy and apoptosis. |
|
|
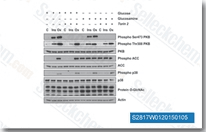
| S2817 | Torin 2 | Torin 2 is a potent and selective mTOR inhibitor with IC50 of 0.25 nM in p53−/− MEFs cell line; 800-fold greater selectivity for mTOR than PI3K and improved pharmacokinetic properties. This compound inhibits ATM/ATR/DNA-PK with EC50 of 28 nM/35 nM/118 nM,in PC3 cell lines respectively. It decreases cell viability and induces autophagy and apoptosis. |
|
|
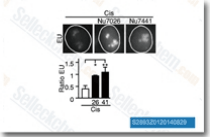
| S2893 | NU7026 | NU7026 (LY293646) is a potent DNA-PK inhibitor with IC50 of 0.23 μM in cell-free assays, 60-fold selective for DNA-PK than PI3K and inactive against both ATM and ATR. This compound enhances G2/M cell arrest and apoptosis. |
|
|
| S7891 | CC-115 | CC-115 is a dual inhibitor of DNA-dependent protein kinase (DNA-PK) and mammalian target of rapamycin (mTOR) with IC50 values of 0.013 μM and 0.021 μM, respectively. It has potential antineoplastic activity. |
|
|
| S8379 | YU238259 | YU238259 is a novel inhibitor of homology-dependent DNA repair(HDR), but does not inhibit non-homologous end-joining (NHEJ), in cell-based GFP reporter assays. |
|
|
Signaling Pathway Map
DNA-dependent protein kinase (DNA-PK) is composed of three key components including two DNA-binding subunits Ku70 and Ku80 (Ku86), as well as one DNA-dependent protein kinase catalytic subunit (DNA-PKcs). Based on protein sequence similarity, DNA-PK belongs to the phosphatidylinositol-3-kinase (PI3K) family, whereas, DNA-PK is not known to phosphorylate lipids and is therefore called PI3K-like kinase (PI3KK). The carboxyl-terminal region of Ku70 contains a SAP domain that is believed to be involved in chromosomal organization. The carboxyl-terminal region of Ku80 is required for the Ku70 and Ku80 heterodimer interaction with DNA-PKcs. The Ku heterodimer can bind to a variety of double-stranded end structures, including blunt ends, overhangs are the 3' or 5' end, and covalently closed hairpin ends. Like ATM and ATR, DNA-PKcs is structurally similar as it contains carboxyl-terminal domains, a large amino-terminal domain in addition to FAT and FATC domains flanking the kinase domain. The DNA-PKcs structure contains a channel large enough to accommodate double-stranded DNA, while the structure of Ku heterodimer is an asymmetric open ring, allowing the DNA to pass through the center. DNA-PKcs is one of the largest kinases identified to date, and it is the only kinase that is absolutely dependent on DNA binding for activity. DNA-PK has a strong preference for phosphorylating Serine (Ser) and Threonine (Thr) residues that are followed by glutamine or, less commonly, a hydrophobic residue. [1][2]
DNA-PK is involved in the ligation step of the non-homologous end joining (NHEJ) pathway required for DNA double-stranded break (DSB) repair, V(D)J recombination and telomere stabilization. A heterodimer of Ku70 and Ku80 initially binds to the double-stranded DNA broken ends and translocates inwards in an ATP-independent manner and recruits DNA-PKcs. This results in the stabilization of the protein/DNA binding and enabling NHEJ to proceed. Moreover, DNA-PKcs acts as a scaffold protein by joining two broken DNA ends together in a complex containing two DNA-PKcs molecules that contributes to the synapsis of the broken DNA ends and the localization of DNA repair proteins such as DNA ligase IV/XRCC4 complex to the site of damage. DNA-PK is activated in cis by the DNA to which it is bound, and stimulated by Ku heterodimer as well as the interaction of two molecules of DNA-PKcs, while end-bridging through synapsis is required for full kinase activation. DNA-PKcs autophosphorylation at multiple sites, including Thr2609 and Ser2056, results in an inactivation of DNA-PK kinase activity and NHEJ ability. To ensure NHEJ can proceed efficiently, DNA-PK phosphorylates and activates the Werner syndrome protein (WRN) to remove 3' phosphate or 3' phosphoglycolate groups generated following IR, and the nuclease Artemis to remove 5' overhangs and shorten 3' overhangs. In addition, DNA-PK promotes processing of hairpin DNA structures in V(D)J recombination by activation of Artemis. Cells that lack DNA-PKcs are acutely radiosensitive and have defective DSB repair, while mice lacking DNA-PKcs remain viable but are immunodeficient (due to the absence of immune development) as a result of accumulated processed DNA intermediates. Additionally, DNA-PK has been strongly implicated in telomere maintenance. DNA-PKcs-/- mice display significant telomeric fusion events consistent with the role of DNA-PKcs in telomere maintenance. Furthermore, DNA-PK is involved in the modulation of transcription by phosphorylation of RNA polymerases including pol I and pol II through its kinase activity, thereby regulating the function of these enzymes. By direct p53 phosphorylation, the modification of Ku70 releasing Bax, or suppressing the expression of p21, DNA-PK plays a significant role in mediating a p53-dependent apoptotic response under a range of cellular conditions including exposure to ionizing radiation (IR), environmental carcinogens and chemotherapeutic agents or in cells that have critically shortened telomeres. [1][2]
Specific inhibitors of DNA-PK used to selectively reduce NHEJ activity have been shown to be effective as single-agent therapies in homologous recombination (HR) -defective tumors. Treatment with a flavone-based DNA-PK inhibitor IC87361 leads to tumor regression. The inhibitors of DNA-PK such as NU7441 enhance the cytotoxicity of physical and chemical agents, leading to reduced clonogenic survival and cellular proliferation, as well as increased apoptosis, regardless of p53 status. Moreover, DNA-PK inhibitors combined with other DNA-damage response (DDR) inhibitors enhance the therapeutic potential of anticancer agents. [3][4]
Products are for research use only. Not for human use. We do not sell to patients.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.