Vascular Endothelial Growth Factor Receptor (VEGFR) is the receptor of VEGF. VEGFR is involved in cell proliferation, migration, survival and permeability. The VEGFs include five known structurally-related mammalian ligands (VEGFA, VEGFB, VEGFC, VEGFD, and placenta growth factor, PLGF) and there are also three structurally related VEGFRs subtypes (VEGFR1, VEGFR2, and VEGFR3).
-
 Australia
Australia
-
 Austria
Austria
-
 Belgium
Belgium
-
 Brazil
Brazil
-
 Canada
Canada
-
 China
China
-
 Czech Republic
Czech Republic
-
 Denmark
Denmark
-
 Finland
Finland
-
 France
France
-
 Germany
Germany
-
 Greece
Greece
-
 Hong Kong
Hong Kong
-
 Hungary
Hungary
-
 Iceland
Iceland
-
 India
India
-
 Ireland
Ireland
-
 Israel
Israel
-
 Italy
Italy
-
 Japan
Japan
-
 Korea
Korea
-
 Luxembourg
Luxembourg
-
 Malaysia
Malaysia
-
 Netherlands
Netherlands
-
 New Zealand
New Zealand
-
 Norway
Norway
-
 Poland
Poland
-
 Qatar
Qatar
-
 Romania
Romania
-
 Saudi Arabia
Saudi Arabia
-
 Singapore
Singapore
-
 Spain
Spain
-
 Sweden
Sweden
-
 Switzerland
Switzerland
-
 Taiwan
Taiwan
-
 Turkey
Turkey
-
 United Kingdom
United Kingdom
-
 United States
United States
VEGFR Inhibitors
Other Protein Tyrosine Kinase Related Inhibitors
| Cat.No. | Product Name | Information | Product Use Citations | Product Validations |
|---|---|---|---|---|
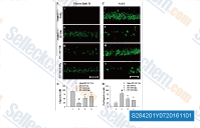
| S2842 | SAR131675 | SAR131675 is a VEGFR3 inhibitor with IC50/Ki of 23 nM/12 nM in cell-free assays, about 50- and 10-fold more selective for VEGFR3 than VEGFR1/2, little activity against Akt1, CDKs, PLK1, EGFR, IGF-1R, c-Met, Flt2 etc. |
|
|
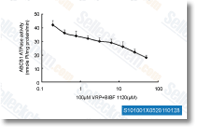
| S1010 | BIBF 1120 (Nintedanib) | Nintedanib is a potent triple angiokinase inhibitor for VEGFR1/2/3, FGFR1/2/3 and PDGFRα/β with IC50 of 34 nM/13 nM/13 nM, 69 nM/37 nM/108 nM and 59 nM/65 nM in cell-free assays. Phase 3. |
|
|
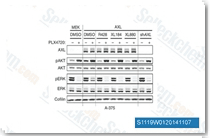
| S1119 | Cabozantinib (XL184) | A potent VEGFR2 inhibitor with IC50 of 0.035 nM, Cabozantinib (XL184) also inhibits c-Met, Ret, Kit, Flt-1/3/4, Tie2, and AXL with IC50 of 1.3 nM, 4 nM, 4.6 nM, 12 nM/11.3 nM/6 nM, 14.3 nM and 7 nM in cell-free assays, respectively. It induces PUMA-dependent apoptosis in colon cancer cells via AKT/GSK-3β/NF-κB signaling pathway. |
|
|
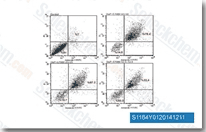
| S1164 | E7080 (Lenvatinib) | Lenvatinib is a multi-target inhibitor, mostly for VEGFR2(KDR)/VEGFR3(Flt-4) with IC50 of 4 nM/5.2 nM, less potent against VEGFR1/Flt-1, ~10-fold more selective for VEGFR2/3 against FGFR1, PDGFRα/β in cell-free assays. Lenvatinib (E7080) also inhibits FGFR1-4, PDGFR, Kit (c-Kit), RET (c-RET), and shows potent antitumor activities. Phase 3. |
|
|
| S1005 | Axitinib (AG-013736) | Axitinib is a multi-target inhibitor of VEGFR1, VEGFR2, VEGFR3, PDGFRβ and c-Kit with IC50 of 0.1 nM, 0.2 nM, 0.1-0.3 nM, 1.6 nM and 1.7 nM in Porcine aorta endothelial cells, respectively. |
|
|
| S7667 | SU 5402 | SU5402 is a potent multi-targeted receptor tyrosine kinase inhibitor with IC50 of 20 nM, 30 nM, and 510 nM for VEGFR2, FGFR1, and PDGF-Rβ, respectively. |
|

|
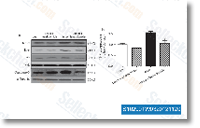
| S8401 | Erdafitinib (JNJ-42756493) | Erdafitinib is a potent and selective orally bioavailable, pan fibroblast growth factor receptor (FGFR) inhibitor with potential antineoplastic activity. This compound also binds to RET (c-RET), CSF-1R, PDGFR-α/PDGFR-β, FLT4, Kit (c-Kit) and VEGFR-2 and induces cellular apoptosis. |
|

|
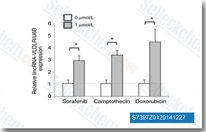
| S7397 | Sorafenib (BAY 43-9006) | Sorafenib is a multikinase inhibitor of Raf-1 and B-Raf with IC50 of 6 nM and 22 nM in cell-free assays, respectively. Sorafenib inhibits VEGFR-2, VEGFR-3, PDGFR-β, Flt-3 and c-KIT with IC50 of 90 nM, 20 nM, 57 nM, 59 nM and 68 nM, respectively. Sorafenib induces autophagy and apoptosis and activates ferroptosis with anti-tumor activity. |
|
|
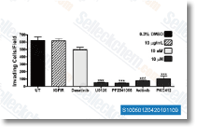
| S1029 | CC-5013 (Lenalidomide) | Lenalidomide is a TNF-α secretion inhibitor with IC50 of 13 nM in PBMCs. Lenalidomide (CC-5013) is a ligand of ubiquitin E3 ligase cereblon (CRBN), and it causes selective ubiquitination and degradation of two lymphoid transcription factors, IKZF1 and IKZF3, by the CRBN-CRL4 ubiquitin ligase. Lenalidomide promotes cleaved caspase-3 expression and inhibit VEGF expression and induces apoptosis. |
|
|
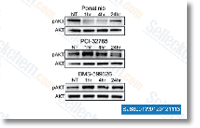
| S1490 | Ponatinib (AP24534) | Ponatinib is a novel, potent multi-target inhibitor of Abl, PDGFRα, VEGFR2, FGFR1 and Src with IC50 of 0.37 nM, 1.1 nM, 1.5 nM, 2.2 nM and 5.4 nM in cell-free assays, respectively. Ponatinib (AP24534) inhibits autophagy. |
|
|
Signaling Pathway Map
Solid tumors require the growth and dissemination of blood vessels and lymphatic vessels to support the metastatic growth of cancers. Following the recognition of growth factor receptor pathways that regulate angiogenesis, a number of small molecular inhibitors and antibodies have been developed that target the activity of vascular endothelial growth factor (VEGF)-VEGF receptor (VEGFR) pathway. This includes oral small-molecule tyrosine kinase inhibitors currently in clinical practice, namely sunitinib and sorafenib. These are commonly used in the treatment algorithm for renal cell carcinoma (RCC) and hepatocellular carcinoma (HCC), two indications that are known to develop resistance to conventional chemotherapeutics.
The VEGFs include five known structurally-related mammalian ligands (VEGFA, VEGFB, VEGFC, VEGFD, and placenta growth factor, PLGF). The VEGFs are disulfide-bonded homodimers, however, VEGFA and PLGF heterodimers are also known to exist. Due to alternative splicing or due to processing, VEGF ligands occur as several different variants. As a result, these variants bind differently to both VEGFRs and to co-receptors resulting in different biological responses including angiogenesis, lymphangiogenesis, permeability, inflammatory cell recruitment and fatty acid uptake. VEGFs are produced by several different cell types and act in a paracrine manner. The VEGFs bind to three structurally related tyrosine kinases (VEGFR1, VEGFR2, and VEGFR3). Modulating the effect of the VEGFRs are a number of co-receptors that lack intrinsic catalytic activity (i.e. heparin sulfate, neurophilins and integrins) and bind to VEGF.[1]
VEGFR1 (also known as Fms-like tyrosine kinase 1, Flt1, in mice) is a single-transmembrane glycoprotein structurally related to VEGFR2 and VEGFR3. VEGFR1 is expressed at high levels in vascular endothelial cells, and along with VEGFR2 binds to VEGFA. VEGFR1 is noted to bind exclusively to VEGFB and PIGF. Expression of VEGFR1 is noted to occur during vessel growth and remodeling activity. Non-endothelial cells that express VEGFR1 includes monocytes and macrophages, human tropholblasts, renal mesangial cells, vascular smooth muscle cells, dendritic cells and various tumor cells. A key regulator of VEGFR1 gene expression is hypoxia.[1]
VEGFR2 (also known as KDR; kinase insert domain receptor, in the human and Flk1; fetal liver kinase-1, in mice) binds VEGFA with a 10-fold lower affinity than VEGFR1. Other targets of VEGFR2 include proteolytically processed VEGFC and VEGFD. The only known ligand to uniquely bind to VEGFR2 is the open reading frame-encoded VEGFE. VEGFR2 is expressed in most adult vascular endothelial cells as well as circulating endothelial progenitor cells, pancreatic duct cells, retinal progenitor cells, megakaryocytes and hematopoietic cells. VEGFR2 expression is induced in conjunction with active angiogenesis (i.e. the uterus during the reproductive cycle) and in pathological process related to neovascularization (i.e. cancer). VEGFR2, often in combination with VEGFR3, is expressed at significantly upregulated levels in the tumor vascular endothelium in most common human solid tumors. Tumor cells can also express VEGFR2, however, epithelial and mesenchymal tumor cells typically express VEGFR1 rather than VEGFR2. Nevertheless, increased expression of VEGFR2 on tumor cells has been noted for melanoma and hematological malignancies. And, there is evidence supporting a relationship between chronic inflammation and tumor development.[1]
VEGFR3 (also known as Fms-like tyrosine kinase 4, Flt4 in the mouse) is activated by the binding of VEGFC or VEGFD, once these two ligands undergo proteolytic processing (this increases their affinity to VEGFR2 and VEGFR3). In addition, hVEGFD shows similar affinity to both VEGFR2 and VEGFR3, while mVEGFD binds only to VEGFR3. During embryogenesis, VEGFR3 expression occurs in the primary vascular plexus at day E8.5. In late stages of embryogenesis, VEGFR3 is expressed in venous endothelial cells of the cardinal vein, that results in VEGFR3-expressing lymphatics. Postnatally, VEGFR3 plays an important role in lymphatic endothelial cells, but its expression is also observed in endothelial cells engaged in active angiogenesis, such as tumor vessels, in endothelial tip cells of angiogenic sprouts in the developing retina or in chronic inflammatory wounds. The receptor is also found in non-endothelial cells such as osteoblasts, neuronal progenitors and macrophages – all of which may indirectly support angiogenesis. It remains unclear if tumor cells express VEGFR3. Despite this lack of clarity, inhibiting VEGFR3 activity is associated with the arrest of tumor vascularization, resulting in decreased vascular density in several tumor models.[1]
Since the VEGF-VEGFR pathway plays a significant role in angiogenesis, and it is widely known that VEGF is highly expressed in tumor and stromal cells, especially in the inflammatory cells of human tumors, dozens of angiogenesis inhibitors are currently undergoing clinical trials.[2] However, despite the number of compounds that has been identified for targeting the VEGF-VEGFR pathway, there is a high attrition rate. Several challenges in the development of angiogenesis inhibitors relate to their specificity, efficacy, side effects, and resistance to anti-angiogenic tumor therapy. However, the emergence of personalized medicine – based on the use of biomarkers – will likely lead to the identification of patient populations that is likely to define respondent groups.
Products are for research use only. Not for human use. We do not sell to patients.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.