-
 Australia
Australia
-
 Austria
Austria
-
 Belgium
Belgium
-
 Brazil
Brazil
-
 Canada
Canada
-
 China
China
-
 Czech Republic
Czech Republic
-
 Denmark
Denmark
-
 Finland
Finland
-
 France
France
-
 Germany
Germany
-
 Greece
Greece
-
 Hong Kong
Hong Kong
-
 Hungary
Hungary
-
 Iceland
Iceland
-
 India
India
-
 Ireland
Ireland
-
 Israel
Israel
-
 Italy
Italy
-
 Japan
Japan
-
 Korea
Korea
-
 Luxembourg
Luxembourg
-
 Malaysia
Malaysia
-
 Netherlands
Netherlands
-
 New Zealand
New Zealand
-
 Norway
Norway
-
 Poland
Poland
-
 Qatar
Qatar
-
 Romania
Romania
-
 Saudi Arabia
Saudi Arabia
-
 Singapore
Singapore
-
 Spain
Spain
-
 Sweden
Sweden
-
 Switzerland
Switzerland
-
 Taiwan
Taiwan
-
 Turkey
Turkey
-
 United Kingdom
United Kingdom
-
 United States
United States
research use only
RALA Antibody (Rabbit mAb) [G17M6]
Cat.No.: F5011
Application:
Reactivity:
-
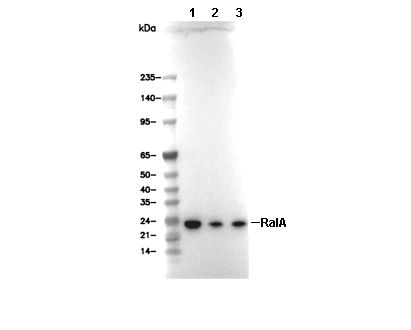 Lane 1: A172, Lane 2: COS-7, Lane 3: Neuro-2a
Lane 1: A172, Lane 2: COS-7, Lane 3: Neuro-2a
Experiment Essentials
WB
Recommended wet transfer conditions: 200 mA, 60 min.
Recommended wet transfer conditions: 200 mA, 60 min.
Usage Information
| Dilution |
|---|
|
| Application |
|---|
| WB |
| Reactivity |
|---|
| Human, Mouse, Rat, Monkey |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW |
|---|
| 24 kDa |
| Positive Control | A172 cells; COS-7 cells; Neuro-2A cells |
|---|---|
| Negative Control |
Experimental Methods
| WB |
|---|
Experimental Protocol:
Sample preparation
1. Tissue: Lyse the tissue sample by adding an appropriate volume of ice-cold RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail),and homogenize the tissue at a low temperature or lyse it by sonication on ice, then incubate on ice for 30 minutes. 2. Adherent cell: Aspirate the culture medium and wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 3. Suspension cell: Transfer the culture medium to a pre-cooled centrifuge tube. Centrifuge and aspirate the supernatant. Wash the cells with ice-cold PBS twice. Lyse the cells by adding an appropriate volume of RIPA/NP-40 Lysis Buffer (containing Protease Inhibitor Cocktail) , sonicate to lyse the cells, and incubate on ice for 30 minutes. 4. Place the lysate into a pre-cooled microcentrifuge tube. Centrifuge at 4°C for 15 min. Collect the supernatant;
5. Remove a small volume of lysate to determine the protein concentration;
6. Combine the lysate with protein loading buffer. Boil 20 µL sample under 95-100°C for 5 min. Centrifuge for 5 min after cool down on ice.
Electrophoretic separation
1. According to the concentration of extracted protein, load appropriate amount of protein sample and marker onto SDS-PAGE gels for electrophoresis. Recommended separating gel (lower gel) concentration: 10%. Reference Table for Selecting SDS-PAGE Separation Gel Concentrations 2. Power up 80V for 30 minutes. Then the power supply is adjusted (110 V~150 V), the Marker is observed, and the electrophoresis can be stopped when the indicator band of the predyed protein Marker where the protein is located is properly separated. (Note that the current should not be too large when electrophoresis, too large current (more than 150 mA) will cause the temperature to rise, affecting the result of running glue. If high currents cannot be avoided, an ice bath can be used to cool the bath.)
Transfer membrane
1. Take out the converter, soak the clip and consumables in the pre-cooled converter;
2. Activate PVDF membrane with methanol for 1 min and rinse with transfer buffer;
3. Install it in the order of "black edge of clip - sponge - filter paper - filter paper - glue -PVDF membrane - filter paper - filter paper - sponge - white edge of clip"; 4. The protein was electrotransferred to PVDF membrane. ( 0.45 µm PVDF membrane is recommended ) Reference Table for Selecting PVDF Membrane Pore Size Specifications Recommended conditions for wet transfer: 200 mA, 60 min. ( Note that the transfer conditions can be adjusted according to the protein size. For high-molecular-weight proteins, a higher current and longer transfer time are recommended. However, ensure that the transfer tank remains at a low temperature to prevent gel melting.)
Block
1. After electrotransfer, wash the film with TBST at room temperature for 5 minutes;
2. Incubate the film in the blocking solution for 1 hour at room temperature;
3. Wash the film with TBST for 3 times, 5 minutes each time.
Antibody incubation
1. Use 5% skim milk powder to prepare the primary antibody working liquid (recommended dilution ratio for primary antibody 1:1000), gently shake and incubate with the film at 4°C overnight; 2. Wash the film with TBST 3 times, 5 minutes each time;
3. Add the secondary antibody to the blocking solution and incubate with the film gently at room temperature for 1 hour;
4. After incubation, wash the film with TBST 3 times for 5 minutes each time.
Antibody staining
1. Add the prepared ECL luminescent substrate (or select other color developing substrate according to the second antibody) and mix evenly;
2. Incubate with the film for 1 minute, remove excess substrate (keep the film moist), wrap with plastic film, and expose in the imaging system. |
Biological Description
| Specificity |
|---|
| RALA Antibody (Rabbit mAb) [G17M6] detects endogenous levels of total RALA protein. |
| Subcellular Location |
|---|
| Cell membrane, Membrane, Mitochondrion |
| Uniprot ID |
|---|
| P11233 |
| Clone |
|---|
| G17M6 |
| Synonym(s) |
|---|
| Ras-related protein Ral-A, RALA, RAL |
| Background |
|---|
| RalA is a Ras‑related small GTPase that cycles between GDP‑ and GTP‑bound states and functions as a membrane‑associated molecular switch downstream of Ras and RalGEFs to control exocyst‑dependent vesicle trafficking, cytokinesis, and oncogenic signaling programs that support proliferation, survival, and metastasis in multiple tumor types. The protein shares the conserved GTP‑binding P‑loop, switch I/II regions, and C‑terminal hypervariable tail with lipidation motifs typical of Ras‑superfamily members, which anchor RalA to specific membrane microdomains and allow GTP‑dependent engagement of effectors such as the exocyst components Sec5 and Exo84, RalBP1/RLIP76, and phospholipase D1, thereby coupling nucleotide state to exocytosis, endocytosis, and actin cytoskeletal regulation. In the canonical Ras pathway, active Ras binds and activates RalGEFs (RalGDS, RGL1–3), which in turn load GTP onto RalA; expression of constitutively active RalA enhances the transforming activity of oncogenic Ras and Raf, while dominant‑negative RalA suppresses Ras‑ and Raf‑driven transformation, demonstrating that a Ras–RalGEF–RalA branch operates in parallel to Raf–MEK–ERK to potentiate oncogenic output. RalA activation promotes exocyst assembly at the plasma membrane and centrosomes, regulates polarized delivery of cargo vesicles, and contributes to successful completion of cytokinesis, and RalA‑dependent exocyst signaling also influences receptor recycling and integrin trafficking, providing a mechanistic route by which RalA can control adhesion, migration, and invasion. Genetic and functional studies in human cancer models show that RalA is required for Ras‑induced tumorigenesis: knockdown or inhibition of RalA in Ras‑mutant cancer cells impairs anchorage‑independent growth, tumor formation, and maintenance of established xenografts, whereas loss of RalB has distinct, more prominent effects on survival and autophagy, highlighting isoform‑specific contributions to the oncogenic Ras network. In triple‑negative breast cancer, elevated RalA activity correlates with aggressive clinical behavior, and experimental silencing of RalA reduces primary tumor growth, limits metastatic colonization, and diminishes cancer stem–like cell properties, positioning RalA as a critical driver of progression and metastasis in this subtype and an attractive target for isoform‑selective inhibition. |
| References |
|---|
|
Tech Support
Tel: +1-832-582-8158 Ext:3
If you have any other enquiries, please leave a message.
Products are for research use only. Not for human use. We do not sell to patients.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.