-
 Australia
Australia
-
 Austria
Austria
-
 Belgium
Belgium
-
 Brazil
Brazil
-
 Canada
Canada
-
 China
China
-
 Czech Republic
Czech Republic
-
 Denmark
Denmark
-
 Finland
Finland
-
 France
France
-
 Germany
Germany
-
 Greece
Greece
-
 Hong Kong
Hong Kong
-
 Hungary
Hungary
-
 Iceland
Iceland
-
 India
India
-
 Ireland
Ireland
-
 Israel
Israel
-
 Italy
Italy
-
 Japan
Japan
-
 Korea
Korea
-
 Luxembourg
Luxembourg
-
 Malaysia
Malaysia
-
 Netherlands
Netherlands
-
 New Zealand
New Zealand
-
 Norway
Norway
-
 Poland
Poland
-
 Qatar
Qatar
-
 Romania
Romania
-
 Saudi Arabia
Saudi Arabia
-
 Singapore
Singapore
-
 Spain
Spain
-
 Sweden
Sweden
-
 Switzerland
Switzerland
-
 Taiwan
Taiwan
-
 Turkey
Turkey
-
 United Kingdom
United Kingdom
-
 United States
United States
research use only
Pyrin Antibody [L17B15]
Cat.No.: F2811
Application:
Reactivity:
-
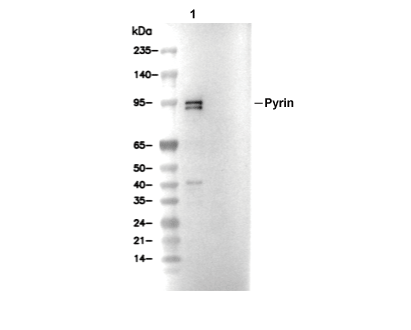 Lane 1: Bone marrow derived-macrophage of WT C57/B6 mice
Lane 1: Bone marrow derived-macrophage of WT C57/B6 mice
Usage Information
| Dilution |
|---|
|
| Application |
|---|
| WB, IP |
| Reactivity |
|---|
| Mouse |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW Observed MW |
|---|
| 86 kDa 110 kDa |
| *Why do the predicted and actual molecular weights differ? The following reasons may explain differences between the predicted and actual protein molecular weight. Post-translational modifications(e.g., phosphorylation, glycosylation); Splice variants and isoforms; Relative charge; Multimerization. |
Biological Description
| Specificity |
|---|
| Pyrin Antibody [L17B15] detects endogenous levels of total Pyrin protein. |
| Clone |
|---|
| L17B15 |
| Synonym(s) |
|---|
| MEF, TRIM20, MEFV, Pyrin, Marenostrin |
| Background |
|---|
| Pyrin is an innate immune sensor encoded by the MEFV gene that belongs to the TRIM family and functions as a cytosolic pattern‑recognition receptor assembling an inflammasome in response to defined disturbances in cytoskeletal and Rho GTPase–dependent homeostasis rather than by directly binding microbial ligands. The protein contains an N‑terminal pyrin domain that mediates homotypic interactions with the adaptor ASC, a central B‑box and coiled‑coil region that support higher‑order assembly and association with cytoskeletal and regulatory partners, and a C‑terminal B30.2/SPRY domain implicated in sensing upstream regulatory changes and in conferring allele‑specific effects on activation threshold. Under resting conditions, RhoA GTPases and their downstream kinases PKN1 and PKN2 phosphorylate pyrin at specific serine residues, creating docking sites for 14‑3‑3 proteins that maintain pyrin in an inactive state, so that intact RhoA signaling keeps the pyrin inflammasome off despite the presence of MEFV. Bacterial toxins and effectors that inactivate RhoA or otherwise disrupt its signaling relieve this repression by preventing pyrin phosphorylation and 14‑3‑3 binding, which permits pyrin oligomerization, ASC recruitment via pyrin–PYD to ASC–PYD interactions, and subsequent recruitment and proximity‑induced activation of pro‑caspase‑1. Activated caspase‑1 cleaves pro–IL‑1β and pro–IL‑18 into their mature forms and processes gasdermin D to drive pyroptotic cell death, thereby coupling detection of pathogen‑induced RhoA inactivation to rapid release of key pro‑inflammatory cytokines and lytic elimination of the infected cell. Pyrin is expressed at high levels in neutrophils, monocytes, and macrophages and contributes to host defense against pathogens that target Rho GTPases, with pyrin‑dependent inflammasome activation lowering bacterial loads and shaping inflammatory responses in lung, gut, and systemic infection models. Missense and truncating mutations in MEFV, particularly within the B30.2 domain, lower the activation threshold or disrupt negative regulation of pyrin, resulting in constitutive or exaggerated inflammasome assembly, increased caspase‑1 activity, and excessive IL‑1β/IL‑18 production that underlie familial Mediterranean fever and other pyrin‑associated autoinflammatory diseases. |
| References |
|---|
|
Tech Support
Tel: +1-832-582-8158 Ext:3
If you have any other enquiries, please leave a message.
Products are for research use only. Not for human use. We do not sell to patients.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.