-
 Australia
Australia
-
 Austria
Austria
-
 Belgium
Belgium
-
 Brazil
Brazil
-
 Canada
Canada
-
 China
China
-
 Czech Republic
Czech Republic
-
 Denmark
Denmark
-
 Finland
Finland
-
 France
France
-
 Germany
Germany
-
 Greece
Greece
-
 Hong Kong
Hong Kong
-
 Hungary
Hungary
-
 Iceland
Iceland
-
 India
India
-
 Ireland
Ireland
-
 Israel
Israel
-
 Italy
Italy
-
 Japan
Japan
-
 Korea
Korea
-
 Luxembourg
Luxembourg
-
 Malaysia
Malaysia
-
 Netherlands
Netherlands
-
 New Zealand
New Zealand
-
 Norway
Norway
-
 Poland
Poland
-
 Qatar
Qatar
-
 Romania
Romania
-
 Saudi Arabia
Saudi Arabia
-
 Singapore
Singapore
-
 Spain
Spain
-
 Sweden
Sweden
-
 Switzerland
Switzerland
-
 Taiwan
Taiwan
-
 Turkey
Turkey
-
 United Kingdom
United Kingdom
-
 United States
United States
research use only
CRTC3 Antibody [M6H14]
Cat.No.: F5006
Application:
Reactivity:
-
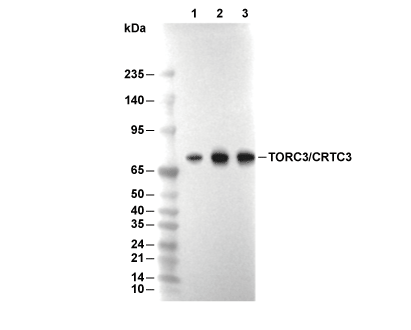 Lane 1: MCF7, Lane 2: Hela, Lane 3: Jurkat
Lane 1: MCF7, Lane 2: Hela, Lane 3: Jurkat
Usage Information
| Dilution |
|---|
|
| Application |
|---|
| WB |
| Reactivity |
|---|
| Human, Mouse, Rat, Monkey |
| Source |
|---|
| Rabbit Monoclonal Antibody |
| Storage Buffer |
|---|
| PBS, pH 7.2+50% Glycerol+0.05% BSA+0.01% NaN3 |
| Storage (from the date of receipt) |
|---|
| -20°C (avoid freeze-thaw cycles), 2 years |
| Predicted MW Observed MW |
|---|
| 70 kDa 76-78 kDa |
| *Why do the predicted and actual molecular weights differ? The following reasons may explain differences between the predicted and actual protein molecular weight. Post-translational modifications(e.g., phosphorylation, glycosylation); Splice variants and isoforms; Relative charge; Multimerization. |
Biological Description
| Specificity |
|---|
| CRTC3 Antibody [M6H14] detects endogenous levels of total CRTC3 protein. |
| Clone |
|---|
| M6H14 |
| Synonym(s) |
|---|
| CREB regulated transcription coactivator 3; CREB-regulated transcription coactivator 3; CRTC3; FLJ21868; TORC-3; TORC3 |
| Background |
|---|
| CRTC3 (CREB‑regulated transcription coactivator 3) is a member of the CRTC family of CREB coactivators that links cAMP and Ca²⁺/stress signaling to transcriptional control of energy metabolism, catecholamine responsiveness, and inflammatory outputs, acting in a phosphorylation‑regulated, Ser133‑independent manner to potentiate CREB1 on both consensus and variant cAMP response elements. The protein contains an N‑terminal CREB‑binding domain with TORC‑conserved regions that dock onto the bZIP region of CREB1 and enhance its interaction with the TFIID subunit TAF4, and a C‑terminal serine‑rich transactivation domain that is heavily phosphorylated by salt‑inducible kinases (SIKs) and other kinases; phosphorylation promotes 14‑3‑3 binding and cytoplasmic retention, whereas dephosphorylation, often via PP2A‑B55 holoenzymes or phosphatases activated downstream of cAMP, calcium, or mitogens, releases CRTC3 to accumulate in the nucleus and coactivate CREB target genes. In adipocytes, CRTC3 is activated by catecholamine signaling through β‑adrenergic receptors in a manner that paradoxically dampens cAMP production: nuclear CRTC3 upregulates the regulator of G‑protein signaling RGS2, which terminates Gs signaling at the β‑adrenergic receptor level and attenuates adenylate cyclase activity, leading to reduced protein kinase A activation, blunted lipolysis, and suppression of thermogenic programs; this negative feedback positions CRTC3 as a transcriptional brake on catecholamine‑driven energy expenditure that promotes positive energy balance and susceptibility to obesity and insulin resistance. Human genetic analyses identify a gain‑of‑function CRTC3 variant associated with increased adiposity, and adipose‑tissue CRTC3 activity increases with high‑fat feeding and obesity, linking this coactivator to metabolic syndrome and making it a potential therapeutic target for enhancing brown and beige fat thermogenesis and improving systemic glucose homeostasis. Beyond adipocytes, CRTC3 acts as a selective coactivator for a subset of CREB‑dependent genes in multiple tissues: it cooperates with PPARGC1A to induce mitochondrial biogenesis in muscle cells, regulates steroidogenic genes such as StAR in endocrine contexts, and functions as a coactivator for HTLV‑1 Tax‑dependent transcription of viral long terminal repeats, integrating hormonal, metabolic, and viral signals at CREB‑controlled promoters and enhancers. In the immune system, phosphorylation of CRTC3 by SIKs in macrophages controls the interconversion between classically activated and regulatory phenotypes, with dephosphorylated CRTC3 entering the nucleus to enhance CREB‑driven IL‑10 production and modulate inflammatory tone, defining a SIK–CRTC3 axis that couples metabolic and stress cues to cytokine programming. |
| References |
|---|
|
Tech Support
Tel: +1-832-582-8158 Ext:3
If you have any other enquiries, please leave a message.
Products are for research use only. Not for human use. We do not sell to patients.
©Copyright 2013 Selleck Chemicals. All Rights Reserved.